Dermatitis de contacto
MODERADOR:
Dra. Mª Mar Garcés.
Hospital Clínico Universitario. Zaragoza.
Bases inmunopatológicas de la dermatitis alérgica de contacto.
Dra. Mª Luisa Sanz
Clínica Universitaria de Navarra. Pamplona.
Resumen
Los mecanismos inmunológicos implicados en DCA incluyen la
participación de diferentes células linfoides y no linfoides de la piel
y de los ganglios regionales. La comunicación entre las diferentes
células puede ser directa o bien mediada por citocinas. El hapteno se
acopla a proteínas plasmáticas del huésped o bien a proteínas
pertenecientes a la propia célula presentadora.
Los mecanismos de activación de respuesta inmune difieren dependiendo
si se trata de una dermatitis de contacto irritativa o alérgica.
Las células de Langerhans (CL) juegan un papel importante en la
presentación del antígeno a la célula T. Los linfocitos T activados por
el antígeno expresan marcadores entre los que se encuentra el antígeno
linfocitario cutáneo CLA+, que confiere a la célula T la propiedad de
migrar desde el ganglio linfático regional, donde se ha activado por
acción de la CL, a la piel, donde ha tenido lugar el contacto
epicutáneo con el hapteno. Esta reacción guarda memoria inmunológica.
Es una enfermedad en cuya patogénesis intervienen numerosos genes,
entre ellos se han detectado polimorfismos en el gen que codifica el
TNF-α.
En el control de la reacción intervienen células T reguladoras y
citocinas como la IL10 y el TGFβ
Definición de dermatitis de contacto e impacto socioeconómico.
La dermatitis de contacto (DC) es una inflamación
dermo-epidérmica, que ocurre tras la exposición de la piel a sustancias
diversas, naturales o sintéticas, cuyo cuadro clínico corresponde al
eczema y a una reacción tipo IV.
Josef Jadassohn describió esta entidad en 1895 y desarrolló la prueba
epicutánea para identificar las sustancias responsables (1).
Constituye una de las más comunes enfermedades de la piel con una
prevalencia estimada del 3% y con un gran impacto socioeconómico, dado
que es la enfermedad ocupacional más frecuente y una de las
enfermedades cutáneas adquiridas más comunes en personas adultas.
Representa entre un 30% y un 40% de todas las enfermedades
ocupacionales y el 90% de las enfermedades cutáneas ocupacionales (2).
Clasificación de la DC según mecanismo
Según el mecanismo patológico implicado se diferencian dos
entidades bien definidas: la DC irritativa, producida por efecto tóxico
e inflamatorio de productos xenobióticos; y la DC alérgica, debida a
una activación T específica (3) (tabla1).
|
Clasificación |
% reacciones |
Agente causal |
Mecanismo |
|
DC alérgica |
20% |
Alérgeno |
Reacción mediada por linfocitos frente a un
hapteno químicamente reactivo. Unido a proteína del paciente. |
|
DC irritativa |
80% |
Irritante
|
Daño tisular directo causado por un irritante. La reacción tóxica depende de la concentración
del irritante |
Dermatitis por contacto causada por irritantes
En este cuadro la sustancia irritante entra en contacto directo
con los queratinocitos de la epidermis. La reacción implica en su fase
final la liberación de citocinas a partir de linfocitos T activados
CD4+ "inespecíficamente" que van a producir IFNa y IL2.
Esta reacción no tiene que ver con haptenos, sino que la inflamación es
subsecuente y directamente proporcional a la concentración de la
sustancia.
Muchas sustancias tanto alergénicas como no alergénicas pueden ser
irritantes a altas concentraciones. La reacción causa la lesión directa
de los tejidos. No requiere memoria inmunológica ni sensibilización
previa (4).
La respuesta inflamatoria inespecífica que interviene en este cuadro
corresponde a la inmunidad innata o no adaptativa, en la que participan
los siguientes mecanismos:
Vasodilatación (aminas vasoactivas)
Liberación de quimiocinas y citocinas (IL1, TNFa)
Aumento de expresión de moléculas de adhesión
Afluencia al lugar de la reacción de células con
capacidad fagocítica
Activación de células NK
Activación inespecífica de células T
Producción de Citocinas. Vía mediada por Factor nuclear kappa-B
La IL1 y el TNFa (citocinas primarias) se almacenan en piel en
grandes cantidades y son liberadas tras un estímulo irritativo, o de
otro tipo.
Tras la unión de sus receptores estas citocinas activan varias vías de
señalización celular, incluyendo el factor nuclear kappa B. Entre los
genes que regulan este factor en células de piel, los que son centrales
en la iniciación de una respuesta inflamatoria cutánea son los genes de
la E-selectina, quimiocinas, citocinas, defensinas, ICAM-1, y VCAM 1.
Pero las citocinas no son el único medio de inducir respuesta NF kappa
B en piel, sino que también a través de la activación de los receptores
Toll-like (receptores que se parecen a la proteína de la drosofila
Toll) se puede activar la vía de señalización NFkB. Estos receptores
reconocen moléculas derivadas de microorganismos, y culmina su
activación en la traslocación del NF kappa B del núcleo y en la
transcripción de genes importantes en la inflamación cutánea que
conlleva a la producción de citoquinas, quimiocinas y moléculas de
adhesión, que dan lugar a formación de mediadores y a la expresión de
receptores celulares que participan en la inflamación (5).
Dermatitis de contacto alérgica (DCA)
En esta entidad clínica se desarrolla una reacción inflamatoria
mediada por células T. Ocurre en el lugar de contacto con el alérgeno
en individuos sensibilizados y los elementos necesarios para el
desarrollo de la CDA son los haptenos, las células presentadoras de Ag
y las células T- específicas.
Esta reacción es altamente específica y guarda memoria inmunológica (3).
Haptenos: Sustancias productoras de DC.
Las sustancias productoras de DC son de origen y naturaleza
diversa.
Normalmente son sustancias químicas de bajo Pm (500-1000 Da), en
algunas ocasiones prohaptenos que sufren metabolización en la epidermis
y cuyo carrier suelen ser proteínas plasmáticas, o bien de la membrana
celular, e incluso glicoproteínas de la clase II del sistema mayor de
histocompatibilidad.
Estas sustancias se encuentran representadas en la naturaleza, algunas
veces de forma muy ubícua (tabla 2).
* Urushiol (sustancia
oleosa de algunas plantas del género Rhus), hiedra venenosa (Toxicodendron radicans), roble
venenoso (Toxicodendron diversilobum),
mango
* Savias de plantas (Philodendron,
hortensias, crisantemos, bulbos de tulipanes)
* Sulfato de níquel
* Dicromato de potasio
* Formaldehído
* Etilendiamina
* Mercaptobenzotiazol
* Thiuram
* Parafenilendiamina
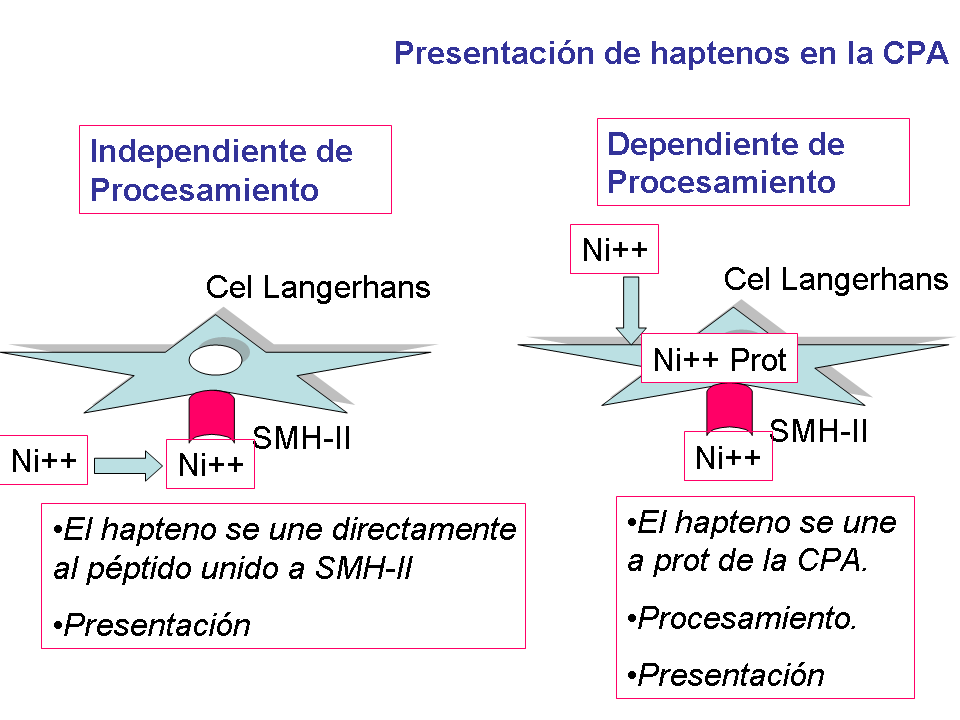
Presentación de haptenos en la célula presentadora de antígeno(CPA)
La presentación de los haptenos al sistema inmune de la piel
puede ocurre mediante dos mecanismos diferentes:
1-Forma clásica: Dependiente de
Procesamiento.
El hapteno se une a una proteína perteneciente a la célula
presentadora de antígeno, tras lo cual se produce el procesamiento del
antígeno por dicha célula y posteriormente la presentación antigénica a
la célula T.
2-. Independiente de Procesamiento.
El hapteno se une directamente al péptido unido a SMH-II.
Posteriormente se produce la presentación al TCR de la célula T (6) (fig1).
Inmunopatología de la DCA
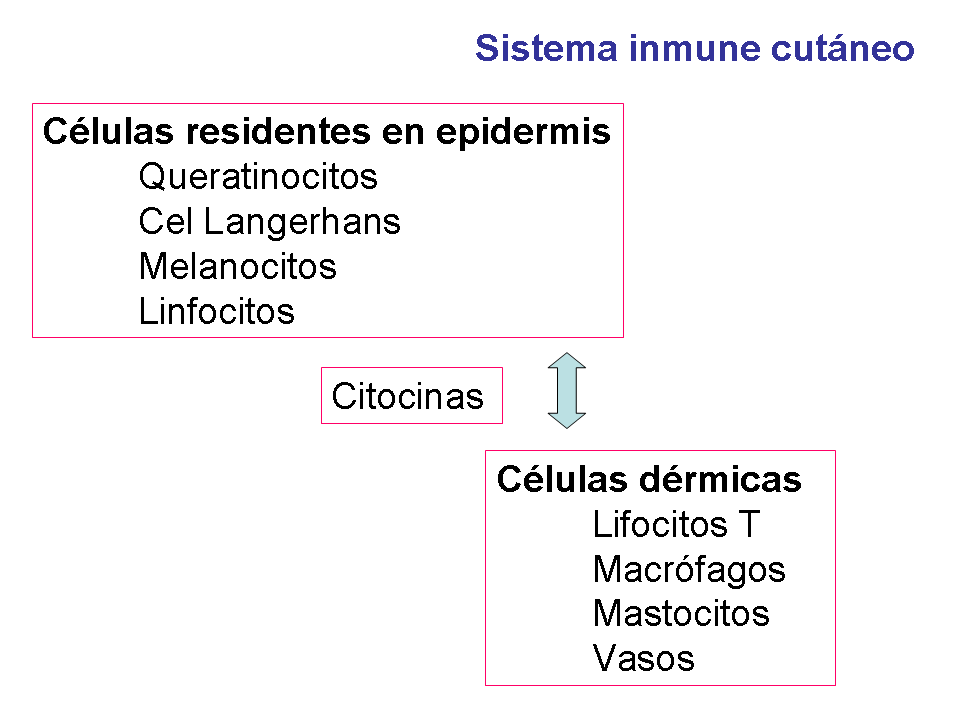
Streilein en 1978 propone la existencia de un sistema
inmunoespecializado que proporciona la inmunidad cutánea e introduce el
concepto de tejido linfoide asociado a piel (SALT) (7).
La respuesta inmune cutánea (tanto innata como adaptativa) involucra la
acción coordinada de las células dérmicas y epidérmicas descritas
anteriormente, mediada por una intrincada red de citocinas (Fig 2).
La dermatitis de contacto alérgica se desarrolla en dos fases bien definidas:
Fase 1: Fase de sensibilización o fase aferente o inductora
Dura desde 10-14 días, hasta semanas tras 1ª exposición al Ag/hapteno.
Cursa asintomática.
Fase 2: Fase de respuesta o fase eferente o fase efectora
Aparece tras horas o días tras la 2ª exposición al Ag. Clínicamente
cursa como una reacción tipo IV (3).
Los acontecimientos celulares ocurren según la siguiente secuencia:
En fase de reposo o no respuesta, las células T que no han tenido
activación previa (células T naive) están recirculando continuamente
entre la sangre y los órganos linfoides, facilitando esta recirculación
la expresión de ciertas proteínas de superficie (L-selectina).
Las células presentadoras de antígeno que están presentes en la piel
-células de Langerhans y células dendríticas dérmicas- captan antígenos
externos y migran a los ganglios linfáticos de drenaje de la piel, por
medio de los vasos linfáticos aferentes, donde presentan el antígeno a
una célula T.
Esta célula T se activa y pasa a ser una célula T de memoria que tiene
como característica especial que expresa CLA (antigeno linfocitario
cutáneo) y un conjunto de citocinas distintivas.
Las células CLA positivas poseen las claves moleculares que les
permiten migrar a la piel, al lugar donde el antígeno inició la
respuesta y algunas de estas células, CLA+, retienen la capacidad de
recircular por el ganglio linfático constituyendo las células T
memoria. (5).
En la dermatitis de contacto alérgica (DAC) el antígeno externo se
introduce epicutáneamente a través de la piel. Los antígenos
sensibilizantes son moléculas reactivas inestables que pueden formar
complejos con las proteínas del huésped.
El antígeno es captado por las células de Langerhans que migran desde
la epidermis hacia los ganglios linfáticos para presentar el antígeno a
la célula T naive. Al cabo de unos días del contacto cutáneo inicial,
las células T de memoria, CLA+, con especificidad para dicho antígeno
salen del ganglio linfático hacia la sangre periférica.
La exposición repetida al antígeno puede aumentar el número de células
T de memoria CLA+ con especificidad para el antígeno hasta alcanzar un
nivel en el cual se produce la dermatitis de contacto. Las células T
CLA+ se extravasan en el lugar de contacto del antígeno, lo reconocen
in situ y se activan. Sus citocinas (y también las procedentes de los
queratinocitos) inducen los cambios inflamatorios que son
característicos de la DAC. Los contactos sucesivos con el antígeno,
incluso meses más tarde repetirán el proceso inflamatorio (5).
Según las fases de la reacción podemos describir los acontecimientos en
varios escalones:
En la fase de sensibilización:
1-Los haptenos penetran en el estrato córneo y son captados por las
células de Langerhans (CL) .
2-Las CL se activan y migran a los ganglios linfáticos .
3-Las CD en migración se localizan en las zonas paracorticales de los
ganglios de drenaje, donde presentan los péptidos-hapteno a las
moléculas clase I y II del SMH de las células T CD4+ y CD8+.
4-Los precursores T específicos se expanden clonalmente en los ganglios
y difunden por la circulación sanguínea a través de los linfáticos
eferentes y el conducto torácico.
Durante este proceso adquieren los Ag de homing (CLA y CCR4) y se
convierten en células T memoria.
Las células T primadas difunden por la piel tras la migración
transendotelial.
Al final de la fase de sensibilización todo está listo para que ocurra
una reacción de dermatitis de contacto cuando entre en contacto la piel
con el hapteno (3).
En la fase de desarrollo de la reacción:
5-Cuando el hapteno entra en contacto por segunda vez, difunde a
través de la epidermis y puede ser cargado por las CL u otras células
de la piel que expresen moléculas del SMH, tales como queratinocitos y
células dendríticas dérmicas que van a ser capaces de activar el
tráfico de células T específicas.
6-La activación de las células T citotóxicas inicia el proceso
inflamatorio que provoca apoptosis de queratinocitos y producción de
citoquinas y quimioquinas.
7-Estas son responsables del reclutamiento de leucocitos (incluidas las
células T reguladoras) de la sangre a la piel y de la aparición de las
lesiones (3).
Importancia de las células T CLA+ en la inflamación cutánea y ligando
del CLA
Durante la fase de sensibilización las células T adquieren los
Ag de homing (CLA y CCR4) y se convierten en células T memoria con el
fenotipo CLA+ y CD45RO+.
Estas células representan el 10-15% cel T sangre y pueden ser CD4 o
CD8, inducen a la formación de citoquinas TH1 y TH2 y median la
inmunovigilancia de la piel.
Las células T CLA+ migran a la piel y recirculan hacia la sangre
durante la inflamación cutánea.
El CLA más que un marcador es una molécula de adhesión que permite la
unión de las cél T al endotelio en las vénulas postcapilares de la piel.
Este paso es necesario y previo a la extravasación de cél T.
La E-selectina es el ligando endotelial del CLA, se expresa en bajos
niveles en los microvasos de piel, y aumenta su expresión en procesos
de inflamación cutánea.
La unión de E-selectina al CLA provoca el acercamiento o rodamiento
hacia pared vascular, la adhesión firme mediada por VCAM1 y otras, y el
paso hacia piel.
Se requiere también en este proceso de activación T la presencia de
quimocinas y firme adhesión de cél T al endotelio a través de
interacciones entre integrinas y moléculas de adhesión.
Cuando estas células llegan a la piel, responden a los Ag y otros
estímulos, producen citocinas y expresan marcadores de activación tales
como el HLA-DR. La IL12, por su parte, induce expresión de CLA en los
linfocitos.
La investigación de estas células CLA+ no sólo es importante para
conocer los mecanismos implicados en esta enfermedad, sino que también
constituye un avance innovador y selectivo para el desarrollo de
posibles nuevos tratamientos para las enfermedades dermatológicas en
general, y para la dermatitis de contacto en particular (5,8,9).
Histopatología de la Dermatitis de contacto alérgica
Los signos histológicos de esta enfermedad más característicos
son los siguientes:
-Espongiosis epidérmica y edema intercelular.
-Epidermotrofismo de céls mononucleares.
-Infiltrado linfohistiocítico.
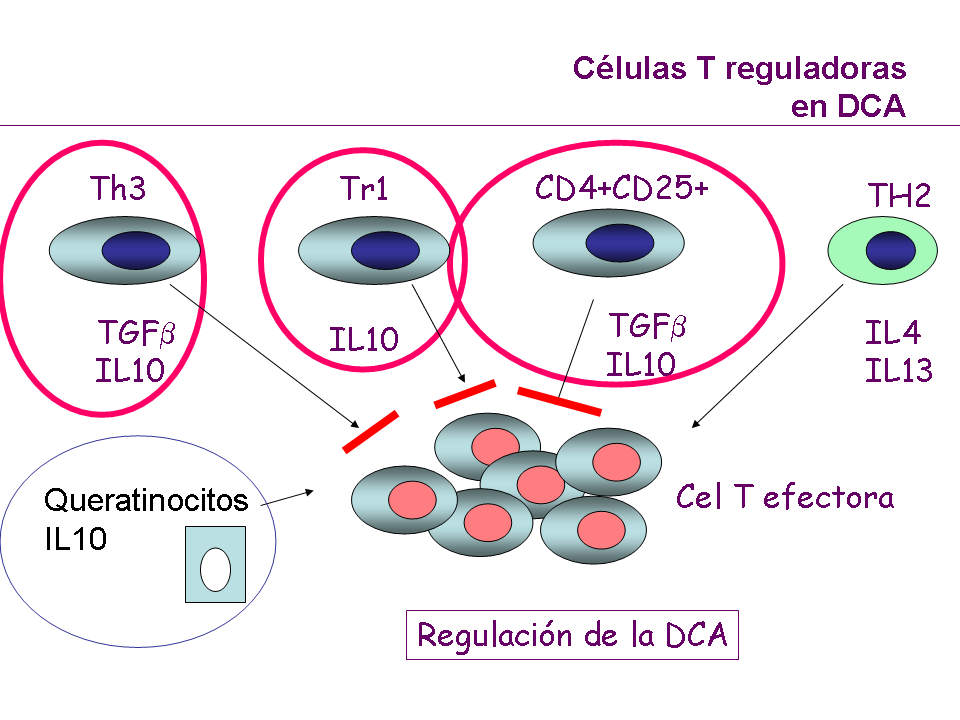
Inmunorregulación de la DCA
Los mecanismos que inciden para limitar la extensión y
perpetuación de las lesiones son: en primer lugar, el aclaramiento del
hapteno en piel, así como la degradación enzimática del Ag, la
destrucción de la CPA, la producción de PGE1 y E2 por parte de los
macrófagos, que serán capaces de inhibir la producción de IL2, citocina
importante en la activación de células T.
Recientemente se ha demostrado que las células T reguladoras CD4 CD25+
y las citocinas reguladoras IL10 y TGFβ son capaces de controlar la
reacción (3,10) (Fig 3).
Papel de las citocinas en la DCA
El concepto clásico de esta enfermedad asume que está causada
por citocinas liberadas de las células Th1 sensibilizadas tras ser
activadas por el Ag (hapteno).
Recientemente se ha comprobado que esto no es así y que además la IL8
parece estar implicada en el mecanismo de acción de la DC irritativa
(11).
En cuanto a la DCA Jensen y cols (12), estudian el perfil de citocinas
y caracterizan las subpoblaciones linfocitarias tras prueba de
provocación oral con níquel, en pacientes con DCA frente a esta
sustancia, y observan que el perfil de citocinas que se produce en
células de sangre periférica corresponde a una producción elevada de
IL5 , lo que corresponde a un patrón TH2 con producción y a una
disminución en sangre periférica de células Cel T memoria (CD3+, CD4+,
CD8+ y CD45RO+), por posible migración a la piel (13).
Por otra parte estudios in vitro realizados por Minang y cols (14)
demuestran que los linfocitos de sangre periférica estimulados con
níquel, (en pacientes alérgicos a esta sustancia) producen citocinas
TH1 (IFNα) y TH2 (IL-4, IL-5 e IL-13), así como la citocina reguladora
IL10.
Estos autores encuentran además una correlación positiva y
significativa entre la producción de citocinas y la intensidad de la
reacción en la prueba epicutánea al níquel.
Polimorfirmos genéticos en la DCA
La patogénesis de la DC implica numerosos genes.
Detectan un polimorfirmo en el gen que codifica el TNF-a, de tal manera que los genotipos
TNFA–308*1/2 y TNFA–308*2/2 tienden a ser más frecuentes entre
individuos polisensibilizados con DC (15).
Diagnóstico in vitro
En cuanto a las técnicas in vitro, se ha demostrado
recientemente que el test de transformación linfoblástico (TTL) es una
técnica fiable en el diagnóstico de la dermatitis de contacto producido
por minoxidil (16).
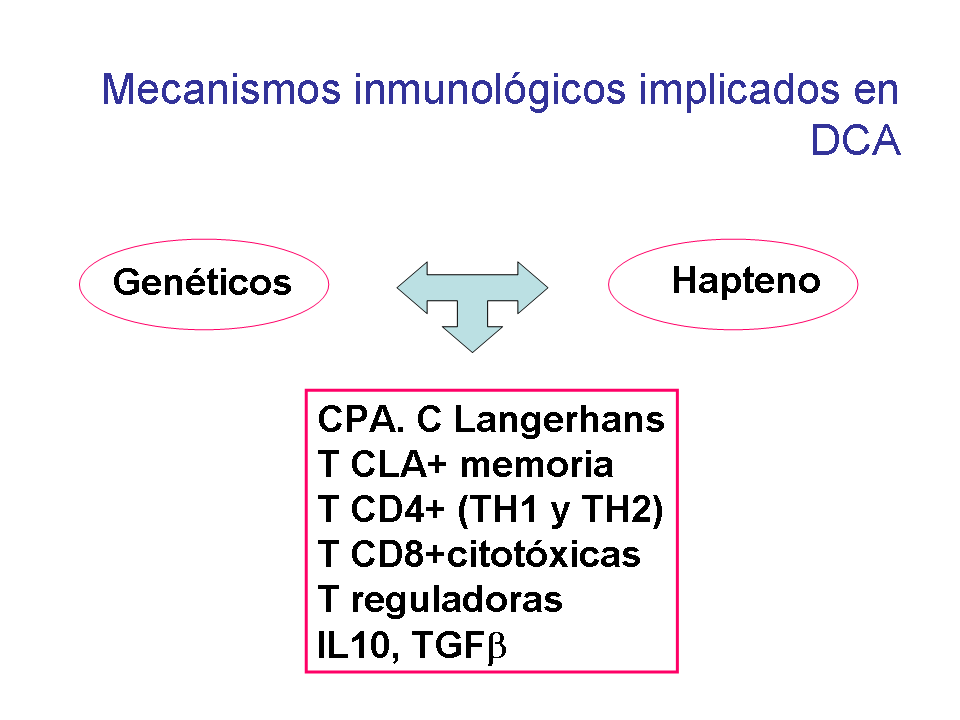
En resumen, la dermatitis de contacto es una enfermedad frecuente, en
la que participa un mecanismo inmune celular, con intervención de
varias células inmunes y no inmunes, como se observa en la figura 4, en respuesta a un hapteno
que contacta con la piel. Este proceso es regulado por citocinas y
moléculas de adhesión, y parece estar controlado genéticamente
Bibliografía
1-Schuler G. Contact allergy Wien Klin Wochenschr. 1993;105:641-7.
Review.
2-Uter W, Schnuch A, Geier J, Frosch PJ. Epidemiology of contact dermatitis. The information network of departments of dermatology in Germany. Eur J Dermatol. 1998; 8: 36-40.
3-Saint-Mezard P, Kastreva M, Berard F, Dubois B, Kaiserlian, D, Nicolas JF. Allergic Contact Dermatitis. En: Skin Immune System Ed JD Bos. CRC Press. Florida USA 2005.
4-Willis CM Variability in responsiveness to irritants: thoughts on possible underlying mechanisms. Contact Dermatitis 2002; 47:267-71.
5-Robert C, Kupper TS. Inflammatory skin diseases, T cells, and immune surveillance. N Engl J Med. 1999 ;9;341:1817-28. Review
6-Emtestam L, Olerup O. On T-cell recognition of nickel as a hapten. Acta Derm Venereol. 1996;76:344-7.
7-Streilein JW, Alard P, Niizeki H. A new concept of skin-associated lymphoid tissue (SALT): UVB light impaired cutaneous immunity reveals a prominent role for cutaneous nerves. Keio J Med. 1999 ;48:22-7. Review.
8- Santamaría-Babi LF. Skin-homing T cells in cutaneous allergic inflammation. Chem Immunol Allergy. 2006;91:87-97.
9-Akdis CA, Akdis M, Simon HU, Blaser K. Regulation of allergic inflammation by skin-homing T cells in allergic eczema. Int Arch Allergy Immunol. 1999;118:140-4. Review.
10-Cavani A, Nasorri F, Ottaviani C, Sebastiani S, De Pita O, Girolomoni G. Human CD25+ regulatory T cells maintain immune tolerance to nickel in healthy, nonallergic individuals. J Immunol. 2003;171:5760-8.
11-Allen DG, Riviere JE, Monteiro-Rivielle NA. Analysis of interleukin-8 release from normal human epidermal keratinocytes exposed to aliphatic hydrocarbons: delivery of hydrocarbons to cell cultures via complexation with alpha-cyclodextrin. Toxicol In Vitro. 2001;15:663-9.
12-Jensen CS, Lisby S, Larsen JK, Veien NK, Menne T.Characterization of lymphocyte subpopulations and cytokine profiles in peripheral blood of nickel-sensitive individuals with systemic contact dermatitis after oral nickel exposure. Contact Dermatitis. 2004;50:31-8.
13-Jensen CS, Menne T, Duus Johansen J. Systemic contact dermatitis after oral exposure to nickel: a review with a modified meta-analysis. Contact Dermatitis. 2006;54:79-86.
14-Minanag JT, Trove-Blomberg M,Lundeberg L, Ahlborg N. Nickel elicits concomitant and correlated in vitro production of Th1-, Th2-type and regulatory cytokines in subjects with contact allergy to nickel. Scand J Immunol. 2005;62:289-96.
15-Westphal GA, Schnuch A, Moessner R, Konig IR, Kranke B, Hallier E, Ziegler A, Reich K. Cytokine gene polymorphisms in allergic contact dermatitis. Contact Dermatitis. 2003;48:93-8.
16-Hagemann T, Schlutter-Bohmer B, Allam JP, Bieber T, Novak N. Positive lymphocyte transformation test in a patient with allergic contact dermatitis of the scalp after short-term use of topical minoxidil solution. Contact Dermatitis. 2005;53:53-5.
{kind=link}
{kind=link}
{kind=link}
{kind=link}