PRIMERA
PONENCIA:
Polimorfismos genéticos
MODERADOR:
Dr. Carlos Colás Sanz.
Hospital Clínico Universitario.
Zaragoza.
Polimorfismos en asma y rinitis
Dres.
Elena Llanes, Blanca Cárdaba, Fernando Florido, Joaquin Quiralte y Carlos Lahoz.
Hospital Fundación Jiménez Díaz. Madrid.
Introducción
El polen del olivo es una de las principales causas de alergia respiratoria en el Área Mediterránea (1, 2). Aunque está descrita la alta prevalencia de los síntomas nasales y conjuntivales inducidos por O. europaea (3), también puede provocar el aumento de la incidencia de asma inducido por este polen entre Abril y Junio.
Los niveles de polen de O. europaea necesarios para desencadenar los síntomas de rinitis alérgica estacional son extremadamente altos (400 granos/m3) comparados con los 50 granos/m3 necesarios en pacientes sensibilizados a gramíneas (4). En Jaén las concentraciones de polen en ambiente oscilan entre 500 y 1000 granos/m3 durante al menos la mitad de la estación polínica, con picos de más de 5000 granos/m3 a mediados de Mayo. Por esta razón, se considera que los pacientes tienen rinitis o asma alérgica más severa cuando están expuestos a estos elevados niveles de polen (5). De hecho, hay evidencias que sugieren una respuesta más compleja de anticuerpos IgE contra los alérgenos del polen del olivo en pacientes alérgicos de Jaén, en condiciones de exposición a niveles elevados de polen, comparados con aquellos pacientes polínicos de otras regiones con exposición a menores niveles de polen (6).
Mediante diferentes técnicas de laboratorio, el extracto del polen del olivo ha mostrado la presencia de 20 proteínas con actividad alergénica (7). Entre ellos, Ole e 1 es el alérgeno de más frecuente sensibilización y es reconocido por más del 70% de los pacientes (8, 9). Junto a Ole e 1, se han aislado y purificado otros nueve alérgenos del polen del olivo (10). Dos de estas proteínas alergénicas, Ole e 4 (11) y Ole e 6 (12) no presentan homología funcional con otras proteínas conocidas. Otros alérgenos se corresponden con proteínas que poseen un alto grado de homología de secuencia con proteínas clonadas de diferentes tejidos vegetales, como la superóxido dismutasa (Ole e 5) (11), proteínas de unión a calcio (Ole e 3 y Ole e 8) (13, 14), proteínas de transferencia de lípidos (Ole e 7) (15, 16) y β1-3 glucanasas (Ole e 9) (17).
El mejor conocimiento de este grupo de proteínas alergénicas podría contribuir a mejorar el diagnóstico de la sensibilización al polen del olivo (21) así como permitir la identificación de factores específicos que influyen en el fenotipo clínico de las enfermedades inducidas por O. europaea.
Por otra parte, la influencia genética sobre la respuesta alérgica en esta clase de enfermedades está bien demostrada (22). Sin embargo, el no haber hallado un modelo de herencia sencilla que explique los desórdenes atópicos implica que este tipo de enfermedades son multifactoriales, con la posible interacción de varios genes con múltiples factores ambientales. Hasta la fecha se han identificado un buen número de genes que influyen en la respuesta alérgica (22, 23), siendo la correlación entre la respuesta de anticuerpos IgE específicos contra alérgenos y antígenos HLA de clase II los más estudiados (23). En este sentido, anteriormente habíamos descrito la relación entre antígenos HLA de clase II y la respuesta de anticuerpos IgE a Ole e 1, en dos poblaciones no relacionadas, donde observamos una fuerte asociación entre HLA-DRB1*0701/2 y HLA-DQB1*0201 y esta respuesta (24-26).
Otros genes se han postulado, también, como candidatos a la regulación de la alergia y el asma.
Un incremento de la respuesta inmunológica Th2 y la producción de citocinas tales como IL-13, IL-4, e IL-5 contribuyen a la inducción de alergia y el asma.
Estas citocinas son responsables de la cascada de activación y reclutamiento de esosinófilos, producción de IgE por las células B y el mucus necesarios para el desencadenamiento de la inflamación alérgica. Los genes de la ruta de señalización de IL4 e IL13 juegan un papel central en la regulación de IgE. Ambas interleucinas, IL4 e IL13, comparten la cadena del receptor IL4Ra.
El gen de IL-13 Gen se encuentra en el cromosoma 5q31-q33, una región que está relacionada con fenotipos de asma. SNPs (polimorfismos nucleotídicos simples) que son funcionales en el promotor y en regiones codificantes de IL-13 han sido asociados con asma, hiperrespuesta de las vías respiratorias, atopia y niveles totales de IgE en suero. Dos polimorfismos de IL13 han sido investigados en relación con el asma. Uno se encuentra localizado en la región promotora, en posición –1112, adyacente a la región que ocupa el factor nuclear de células T activadas y ha sido asociado con el asma alérgico en una población alemana y con la regulación alterada de la producción de IL13. Otro es una modificación G – A en el nucleótido +2044 en la región codificante del exón 4 que conduce a la sustitución del aminoácido R130Q. El polimorfismo R130Q ha sido asociado con niveles totales de IgE elevados, dermatitis atópica y asma en varias poblaciones.
El gen que codifica para la cadena IL4Ra (IL4RA) está localizado sobre el cromosoma 16p12.1. Dos polimorfismos han sido identificados en la región codificante del gen de IL4RA, los cuales conducen a un cambio de aminoácido en el receptor. Una variante extracelular de la cadena de IL4Ra consistente en una sustitución del aminoácido valina en posición 50 por isoleucina, ha sido asociado con atopia y asma. Una variante del dominio intracelular de IL4Ra consistente en la sustitución de glutamina en posición 551 por arginina (Q551R) se ha asociado con el síndrome hiper-IgE, eczema atopico severo y aumento de la señalización a través de IL4Ra. El alelo arginina 551 (551R) se asocia con atopia. Estudios funcionales han revelado que el alelo 551R produce un receptor que muestra un aumento de la transducción de señales in vitro. La homocigosidad para el alelo 551R ha sido asociada con asma severo. IL-5 es un importante determinante de la respuesta Th2 response. El gen de IL-5 se encuentra localizado en el cromosoma 5q31.1, y algunos grupos lo han asociado con el asma. La actividad biológica de IL-5 está específicamente dirigida hacia el desarrollo, diferenciación, reclutamiento, activación y supervivencia de los eosinófilos. La inhalación del alérgeno incrementa la producción de IL-5 en las vías respiratorias. También incrementa el número de eosinófilos en sangre periférica y los linfocitos que contienen IL-5 intracelular.
IL-5 C-746T, en la región promotora del gen, es el único polimorfismo de IL-5 descrito en la población europea.
El gen b2Adrenergico está localizado en el cromosoma 5q31-q33. B2AR es una proteína transmembrana expresada por las células del músculo liso de las vías respiratorias. Es responsable de la unión de las catecolaminas endógenas, las cuales median su acción sobre un amplio tipo de células, y han sido implicadas en la patogénesis del asma. Dos polimorfismos en el gen B2AR dan lugar a la sustitución de un aminoácido en el codon 16, con el cambio de Arg por Gly (R16G), y en el codon 27, con Glu en lugar de Gly (Q27E); los cuales son responsables de los cambios en la región extracelular de B2AR. Se sabe que ambos polimorfismos alteran la función del receptor “in vitro”. Con estos antecedentes, nos propusimos:
El estudio de un grupo de 146 pacientes procedentes de Jaen, que sufrían polinosis a O. Europaea y fueron seleccionados por los Drs. Quiralte y Florido, evaluando: Tipo y severidad clínica de la enfermedad y la inmunogenicidad de los alérgenos purificados del polen del olivo usando test cutáneos así como ELISA de anticuerpos IgE frente a los antígenos del olivo utilizados.
Una vez obtenidos los valores clínicos e inmunológicos de nuestra población, decidimos analizar si había algún tipo de implicación clínica relacionada con la reactividad IgE específica de alérgeno, además de la posible interrelación entre estas sensibilizaciones y el perfil genético, empezando por el loci HLA de clase II DR y DQ.
Además queríamos analizar el papel de los polimorfismos: IL13 C–1112T, IL13 R130Q, IL4RA I50V, IL4RA Q551R, IL5 C–746T, B2AR R16G y Q27E, en la respuesta inmune de pacientes alérgicos al polen del olivo. Con este propósito estudiamos la distribución de frecuencias genotípicas entre alérgicos y controles, la distribución de frecuencias entre asmáticos y riníticos, si existían diferencias significativas en la sensibilización específica a los alérgenos del polen del olivo según el genotipo y finalmente si los niveles de IgE (totales y específicos) estaban relacionados con un genotipo en particular.
Material y Métodos
Pacientes y Controles
Para nuestro estudio seleccionamos 146 pacientes no relacionados, reclutados en el Departamento de Alergia del “Hospital Ciudad de Jaén”, con los siguientes criterios: rinitis y/ó asma estacional de Abril a Junio, test cutáneo positivo al extracto del polen de O. europaea (ALK Abelló, Madrid, España), y no haber recibido inmunoterapia previa a O. europaea. Como grupo control reclutamos 50 individuos sanos de la misma área geográfica. Se obtuvo el consentimiento informado de cada uno de los participantes. Así mismo se obtuvo la aprobación del Comité Ético y Científico del “Hospital Ciudad de Jaén” para llevar a cabo en humanos test cutáneos con alérgenos purificados.
Evaluación clínica
El registro de los síntomas de los pacientes, la medicación utilizada y el flujo espiratorio (PEFR) se recogió diariamente de Abril a Junio de 2000. Los síntomas oculares y nasales (estornudos, congestión, enrojecimiento y picor), y los síntomas torácicos (jadeos, asma y congestión) fueron registrados en una escala de 0–3 (una puntuación de 0 indicó ausencia de síntomas y entre 1–3, síntomas leves, moderados, y severos, respectivamente). Los pacientes recogieron en un registro diario los datos de asma o rinitis, las veces que se despertaban a lo largo de la noche y síntomas dudosos de asma, empleo de medicación, y medidas de PEFR mañana y tarde. Los pacientes usaron un Mini-Wright peak flow meter (Clement-Clarke Ltd, Harlow, UK) para medir el PEFR y registrar la exhalación forzada más alta, de tres realizadas, al despertar y por la tarde. Los pacientes recibieron azelastina en gotas oculares y spray nasal, loratadina y salbutamol inhalado, como medicación de rescate. Si los síntomas no eran controlados, a los pacientes se les administraba, además, diariamente durante 7 días 30 mg de prednisona. Las variables clínicas más intensas fueron definidas como asma. Para considerar un día con asma, se debe cumplir al menos alguno de los siguientes criterios: puntuación de los valores de asma de ± 2, pico del flujo espiratorio (PEF) matinal un 20% menor que la media de los PEF de los últimos 7 días antes de la polinización y uso diario de salbutamol una ó más veces por encima del uso medio de los últimos 7 días antes de la polinización.
Alérgenos del polen del Olivo
El polen de Olivo se obtuvo de Allergom AB (Ängelholm, Suecia). El polen (5% peso/volumen) fue extraído con 50 mM bicarbonato amónico, pH 8.0, conteniendo 1 mM fluoruro fenil-metil-sulfonil, seguido de centrifugación a 12 000 g durante 20 min a 4º C. El sobrenadante liofilizado fue almacenado a –20º C. Los distintos alérgenos del polen del olivo fueron aislados por el grupo de la Prof. R. Rodríguez de la Universidad Complutense, como se describe en la bibliografía (18, 20). Los niveles de anticuerpos IgE totales en suero fueron determinados mediante el inmunoensayo enzimático para IgE de Pharmacia (Uppsala, Suecia). Los anticuerpos IgE específicos para el extracto de O. europaea fueron determinados mediante el sistema UNI-CAP de Pharmacia. Los niveles de IgE específicos de alérgeno, en los sujetos alérgicos, se determinaron por ELISA.
Test cutáneo
El test cutáneo con el extracto completo de O. europaea y con los distintos alérgenos purificados se realizó en duplicado por punción con una lanceta estándar de 1 mm sobre el antebrazo, según las indicaciones de la EAACI (27). Empleamos histamina fosfato (10mg/ml) como control positivo y solución salina fisiológica como control negativo. Los diferentes alérgenos fueron analizados por duplicado en cuatro diluciones 10X en solución salina (de 100 a 0.1 mg/ml). Las reacciones se determinaron después de 15 minutos midiendo la longitud máxima (D) y el diámetro transversal (d) de la roncha producida. El área de la reacción se calculó mediante la fórmula: [(D + d)/2]2. Los pacientes se clasificaron como sensibilizados a un alérgeno específico si presentaba un área de roncha de al menos 9 mm2 y niveles de IgE alérgeno específicos detectables (superiores a la media de los niveles de IgE de los controles ± 2 DE)
Tipificación de HLA de clase II (DRB1 y DQB1)
Se extrajo el ADN genómico de sangre periférica siguiendo la técnica de precipitación con sales (28) y se cuantificó por espectrofotometría antes de su uso. Se genotipó el ADN genómico de todos los individuos para los loci de clase II DRB1 y DQB1. Los polimorfismos de los loci DRB1, DRB3, DRB4, DRB5 y DQB1 se determinaron por amplificación con oligonucleótidos específicos de secuencia, siguiendo el protocolo del proveedor (Micro-SSP; One lambda, CA, USA)
Tipificación de Polimorfismos
El ADN genómico de todos los individuos se tipificó para los polimorfismos: IL13 C–1112T, IL13 R130Q, IL4RA I50V, IL4RA Q551R, IL5 C–746T, B2AR R16G y Q27E. Los SNPs estudiados con sus nombres, región de localización, nombre alternativo e identificación dbSNP se muestran en la Tabla I.
Los genotipos para los polimorfismos IL13 C–1112T, IL4RA I50V, B2AR R16G y Q27E se determinaron usando el método de reacción en cadena de la polimerasa – restricción de la longitud de fragmentos (PCR-RFLP). Las reacciones de PCR se realizaron en un volumen total de 15 ml conteniendo 30 ng de ADN genómico, 1.5 mM MgCl2 y 0.5 mM de cada primer. Las condiciones fueron comunes para IL13 C-1112T, IL4RA I50V y B2AR Q27E. Consistentes en un paso inicial de desnaturalización a 94 ºC durante 2 min. seguido de 35 ciclos de 94 ºC durante 60 s, la temperatura específica de alineamiento para cada polimorfismo 60 s, 72 ºC, y un paso de extensión final de 10 min a 72 ºC. El producto de PCR amplificado se digirió con endonucleasas de restricción específicas (New England Biolabs, Ipswich, MA, United States) a 37 ºC 60 min. La reacción de restricción se realizó en un volumen final de 15 ml conteniendo 7 ml de producto de PCR. Los fragmentos obtenidos fueron identificados con ABI PRISM 310 Genetic Analyzer (Applied Biosystems, Foster City, CA, United States).
Los cebadores usados para la detección del polimorfismo C–1112T en el promotor de IL13, el polimorfismo I50V en el exón 5 de IL4RA y el polimorfismo Q27E B2AR fueron descritos previamente (32, 33, 34).
El polimorfismo R16G del exón 1 de B2AR fue tipificado con los primers descritos anteriormente en la bibliografía (35). Las condiciones de carrera fueron: desnaturalización a 94 ºC 2 min, un primer paso de amplificación de10 ciclos a 94 ºC 30 s, 60 ºC 15 s, 72 ºC 30 s, y un segundo paso de amplificación de 20 ciclos a 89 ºC 30 s, 60 ºC 15 s, 72 ºC 30 s, seguido por 72 ºC 10 min. El producto de PCR se digirió añadiendo 2U de NcoI (New England Biolabs, Ipswich, MA, United States) e incubando a 37 ºC 3 horas.
La tipificación de los polimorfismos de IL13 R130Q, IL4RA Q551R y IL5 C–746T se realizó de acuerdo con el protocolo de discriminación alélica de Applied Biosystems (Applied Biosystems 7500 Real Time PCR System). Las muestras se amplificaron de acuerdo a las condiciones recomendadas por Applied Biosystems (Foster City, CA, United States). Después de la amplificación, se detectó la fluorescencia con 7500 Real Time PCR System (Applied Biosystems, Foster City, CA, United States) y los genotipos se asignaron automáticamente mediante el programa SDS proporcionado con el instrumento.
Análisis estadístico
Los análisis estadísticos realizados fueron:
1. La significación estadística de las frecuencias fenotípicas entre los grupos caso y control y fenotipo clínico (asma/rinitis) fue medida mediante los test de Fisher y Chi Cuadrado (χ2).
2. Los test de Cochran y Mantel- Haenszel, y el test de ANOVA se usaron para la comparación estadística de las frecuencias fenotípicas entre los grupos de enfermos sensibilizados y no sensibilizados a los distintos alérgenos de Olea europaea.
3. Análisis de regresión múltiple para determinar la posible correlación entre los distintos polimorfismos y los niveles de anticuerpos IgE total, IgE específica al extracto completo de Olea europaea, e IgE específica frente a los distintos alérgenos de O. europaea.
4. Todos los SNPs fueron inicialmente analizados individualmente creándose las categorías genotípicas. Las frecuencias haplotípicas para todos los polimorfismos fueron estimadas mediante el algoritmo de maximización de la esperanza (E.M) mediante el programa Arlequin (Arlequin v3.01, CMPG, University of Berne). El análisis estadístico de los haplotipos se realizó con el paquete estadístico EpiInfo v5.00 (CDC, Atlanta, USA).
Resultados
Pacientes
Las variables clínicas de los pacientes se resumen en la TABLA IV.
Prevalencia de IgE a los alérgenos del olivo y correlación clínica
Todos los sujetos alérgicos (n: 146) presentaban niveles significativos de anticuerpos IgE específicos al extracto completo de olivo, aunque se observaron diferentes frecuencias de sensibilización a los alérgenos purificados. Un total de 117 (80.1%) pacientes reaccionaron al menos a uno de los dos alérgenos que estudiamos. Los 29 restantes estaban sensibilizados a los otros alérgenos. 102 de los pacientes (69.9%) tenían una respuesta significativa de anticuerpos IgE a Ole e 2 y 79 pacientes (54%) reaccionaban a Ole e 10.
La relación entre la respuesta IgE alérgeno específica y el fenotipo clínico muestra una asociación estadísticamente significativa entre la reactividad a Ole e 2 y el asma (p: 0.04, OR: 2.2, IC: 0.9-5.1) y entre Ole e 10 y asma (p: 0.007, OR: 2.8, CI: 1.3– 6.1). Esta asociación no la encontramos con el resto de alérgenos que analizamos en la misma población. Al final del período de polinización, los pacientes asmáticos Ole e 10 positivos también sufrían, de forma significativa, más días con asma en comparación con aquellos pacientes asmáticos que no presentaban una respuesta de anticuerpos IgE específica frente a este alérgeno en particular (13.4 ± 8.7 días vs 9.2 ± 9.1 días, Mann–Whitney U-test, P < 0.05).
También hemos analizado la relación de varios fenotipos inmunológicos (niveles de IgE total, niveles de anticuerpos IgE α-O. europaea, niveles de anticuerpos IgE α-Ole e 2 y α-Ole e 10) con el tipo de enfermedad inducida por el polen del olivo. Para ello, los pacientes fueron clasificados en cuatro grupos de acuerdo a la reactividad que manifiesten para cada alérgeno (TABLA V): 1) un primer grupo compuesto por aquellos que sólo presentaban reactividad contra Ole e 2 (n:38), 2) el segundo grupo reaccionaba exclusivamente a Ole e 10 (n: 15), 3) el tercer grupo presentaba de manera simultánea reactividad frente a estos dos alérgenos (n: 64), y finalmente 4) el último grupo, estaba integrado por pacientes alérgicos al olivo que no presentaban reactividad a estos dos alérgenos (n: 29). De acuerdo a la respuesta de anticuerpos, encontramos que los niveles de anticuerpos IgE más elevados (total, α-Olea europaea, y alérgeno específico) estuvieron asociados con el fenotipo clínico asmático. Por otro lado, los resultados más significativos los encontramos en el grupo que tenía simultáneamente reactividad a los dos antígenos. Este grupo también presentaba los niveles más altos de anticuerpos y el mayor factor de riesgo, OR: 4.3 (p: 0.002).
Tipificación de HLA de clase II
Para analizar la relación entre sensibilización específica y los antígenos HLA, analizamos la distribución de antígenos HLA de clase II de acuerdo a la clasificación anterior de reactividad IgE a alérgenos. Comparamos estos grupos con un grupo adicional de 50 individuos control, no atópicos, de la misma área geográfica. Este análisis, apuntó hacia cuatro antígenos: DR7 y DQ2, DR2 (15) y DR3. (TABLA VI)
El haplotipo DR7-DQ2 parece ser un factor de riesgo frente a la sensibilización a Ole e 2: en primer lugar, la frecuencia fenotípica de DR7 está aumentada, de forma estadísticamente significativa (p corregida: 0.003, OR: 5.4), comparando el grupo de pacientes sensibilizados a Ole e 2 (FF: 60.5%) con los controles (FF: 22%). En segundo lugar, DQ2 está incrementado de forma estadísticamente significativa en los sensibilizados a Ole e 2 (FF: 68.4%) comparados con los pacientes del grupo con reactividad a ambos alérgenos (FF: 37.5%), siendo el valor de p corregida: 0.02 y OR : 3.6.
DR2 (15) está incrementado significativamente (p corregida: 0.03, OR: 5.6) en el grupo de pacientes sensibilizados a Ole e 2 y Ole e 10 (FF: 32.8%) comparado con el grupo control (FF : 8%) y DR3 está estadísticamente disminuido en ese mismo grupo comparado con el grupo control (FF : 32%) (p corregida 0.02, OR: 0.18)
Tipificación de Polimorfismos
La distribución de genotipos de los 7 polimorfismo estudiados esta en equilibrio de Hardy-Weinberg. Los datos de las frecuencias genotípicas de pacientes y controles, así como de asmáticos y riníticos se muestran en la TABLA VII.
IL13 SNPs
Se tipificaron 2 SNPs del gen de IL13 (C-1112T, R130Q)
C-1112T
Investigamos la posible relación entre el polimorfismo IL13 C-1112T y la susceptibilidad de desarrollar alergia al polen del olivo. La distribución de frecuencias genotípicas de este polimorfismo se muestra en la TABLA VII. Encontramos diferencias estadísticamente significativas en la distribución de la frecuencia del genotipo homocigoto TT entre pacientes y controles (P = .006), los pacientes con este genotipo tienen un menor riesgo de desarrollar alergia al polen del olivo (OR, 0.35; 95% CI, 0.16-0.76). Cuando analizamos la distribución genotípica de este polimorfismo de acuerdo al fenotipo clínico de asma o rinitis, niveles de IgE total, niveles de anticuerpos IgE α-Olea y niveles de anticuerpos IgE α-Ole e 1, e 2, e 3, e 6, e 7, e 8, e 10, no encontramos ninguna asociación.
R130Q
El análisis de la distribución de frecuencias genotípicas del polimorfismo IL13 R130Q muestra un incremento estadísticamente significativo (P = .009) del genotipo heterocigoto RQ en pacientes (37.2%) comparado con el grupo control (16%). Esto datos se encuentran recogidos en la TABLA VII. Los individuos con este genotipo tienen un riesgo 3 veces mayor de desarrollar alergia a O.europaea que aquellos que no lo tienen (OR, 3.12; 95% 1.29-7.79). Sin embargo, no encontramos diferencias estadísticamente significativas entre pacientes con asma o rinitis. Tampoco existe asociación con los niveles de IgE. Por otro lado, observamos un incremento significativo (P = .042) en la prevalencia de individuos con genotipo homocigoto en el grupo de pacientes sensibilizados a Ole e 3 (71.4%; n = 40) comparado con los no sensibilizados a ese alérgeno (52.8%; n = 47).
IL4RA SNPs
I50V
No encontramos diferencias significativas en la distribución genotípica de IL4RA I50V entre casos y controles y asmáticos y riníticos.
Sin embargo, el análisis de los niveles de anticuerpos IgE específicos a los alérgenos de polen del olivo mostró diferencias estadísticamente significativas (P = .018). El genotipo homocigoto I50 estaba disminuido en los pacientes sensibilizados a Ole e 3 (19.3%) comparados con los no sensibilizados a Ole e 3 (38.2%). Este genotipo parece ser un factor de protección frente a la sensibilización específica a Ole e 3. Entre los pacientes con anticuerpos específicos α-O.europaea (n = 128) observamos que el genotipo homocigoto V50 esta asociado (P = .045) con los mayores niveles de IgE α-O.europaea en suero (48.35±8.26 IU/ml) comparado con los homocigotos I50 (38±5.48 IU/ml) y los heterocigotos I50V (32.8±4.18 IU/ml).
Q551R
No encontramos diferencias estadísticamente significativas en la prevalencia de frecuencias genotípicas del polimorfismo IL4RA Q551R comparando alérgicos al polen del olivo con controles y asmáticos con riníticos. El análisis de anticuerpos IgE totales y específicos tampoco reveló ninguna asociación estadísticamente significativa con este polimorfismo.
IL5 SNP
C-746T
La asociación entre la alergia a O.europaea y el polimorfismo IL5 (C-746T) se investigó comparando la ocurrencia del polimorfismo en alérgicos y controles. Como se muestra en la TABLA VII, la distribución de frecuencias genotípicas es similar entre casos y controles. Sin embargo, observamos un descenso del genotipo TT en pacientes con asma (7.3%, n = 8) comparado con aquellos que presentan rinitis (18.9%, n = 7), (P = .045, OR, 0.34 95%, CI 0.1-1.15).
El análisis de correlación entre sensibilización alérgeno específica y distribución genotípica no muestra diferencias estadísticamente significativas. Por otro lado, de acuerdo a la sensibilización específica al alérgeno Ole e 2 los mayores niveles de anticuerpos IgE α-Ole e 2 se encuentran asociados (P = .02) al genotipo TT (1.67±0.31 IU/ml; n = 10) comparado con heterocigotos CT (0.847±0.11 IU/ml; n = 46) y homocigotos CC (0.746±0.11 IU/ml; n = 45)
B2AR SNPs
Determinamos las frecuencias genotípicas de los individuos alérgicos al polen del olivo para los polimorfismos B2AR R16G y B2AR Q27E. Para B2AR R16G la prevalencia de homocigotos R16 fue 16.6% (n = 24), heterocigotos R16G 49.7% (n = 72) y homocigotos G16 33.8% (n = 49). Las frecuencias genotípicas del polimorfismo B2AR Q27E fueron homocigotos Q27 43% (n = 62), heterocigotos Q27E 41% (n = 59) y 16% (n = 23) para los homocigotos E27. No observamos ninguna asociación entre los polimorfismos B2AR R16G y Q27E y el riesgo de desarrollar alergia al polen del olivo. No se encontró relación, tampoco, entre la distribución de frecuencias genotípicas de asmáticos y riníticos y los polimorfismos del gen B2AR en estudio.
Análisis de haplotipos
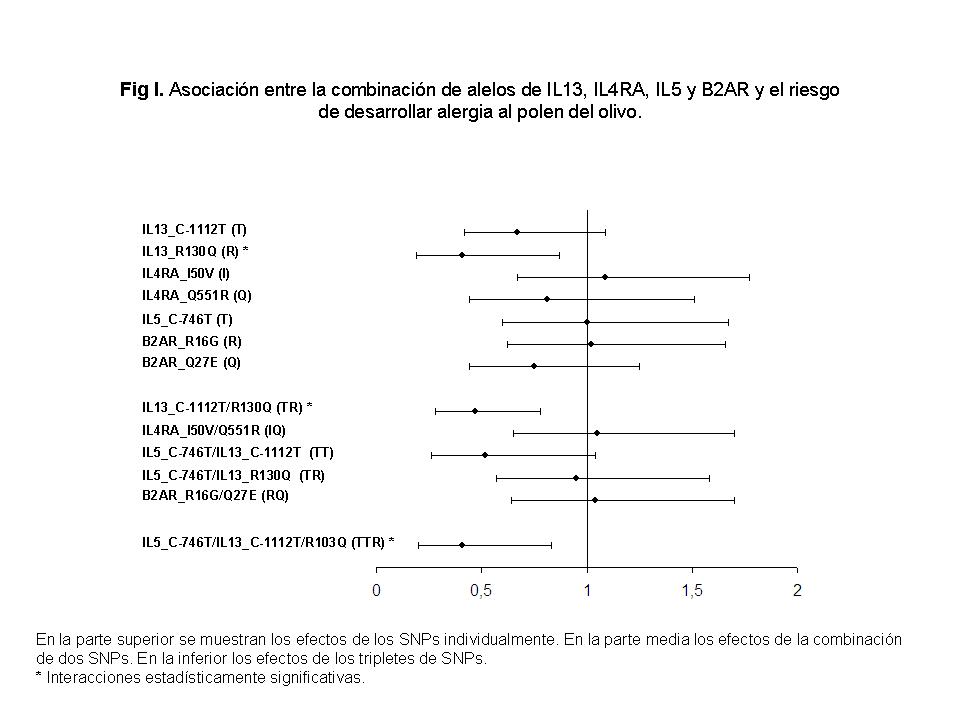
Las interacciones gen-gen también se tuvieron en cuenta. Con el fin de investigar el efecto combinado de los SNPs sobre el desarrollo de la alergia a O.europaea en nuestros pacientes, llevamos a cabo el análisis de haplotipos. Los resultados de este análisis se representan en las Figura I y Figura II.
Los efectos de los alelos de los respectivos SNPs sobre el riesgo de desarrollar alergia a O.europaea se muestran en la parte superior de la Figura I. En un segundo paso, los pares de SNPs localizados sobre el mismo cromosoma fueron analizados, y finalmente, se analizaron en tripletes. De esta manera, el menor riesgo (P = .017) de desarrollar alergia al polen del olivo, para un alelo individualmente, está asociado con el alelo R del polimorfismo IL13 R130Q (OR, 0.41; 95% CI, 0.19-0.87).
Para efectos combinados de SNPs el haplotipo IL13 -1112T/130R tiene un menor riesgo (P = .0025) de alergia (OR, 0,47; 95% CI, 0,28-0,78), así como el triplete formado por IL5 746T/IL13 -1112T/130R (P = .011) (OR, 0,41; 95% CI, 0,2-0,83).
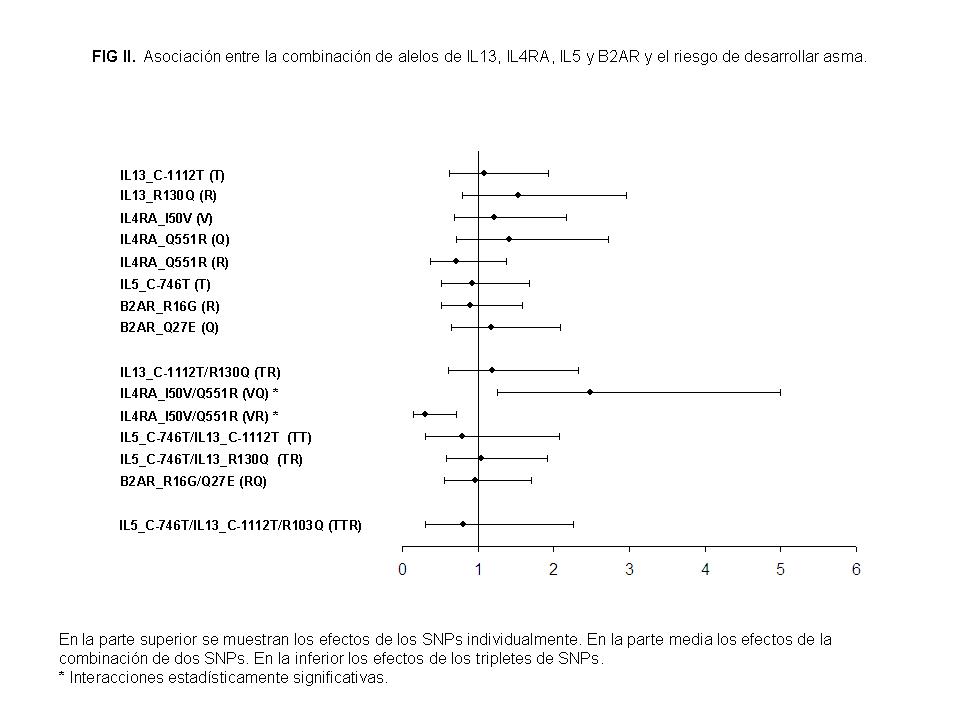
En un segundo análisis, se estudiaron los efectos de la combinación genética sobre el fenotipo clínico (Asma versus Rinitis) de acuerdo al mismo procedimiento. En la Figura II se observa que el OR para cada polimorfismo individualmente es similar entre asmáticos y no asmáticos. Sin embargo, observamos una interacción entre los dos polimorfismos analizados de IL4RA. Encontramos un OR estadísticamente significativo para el haplotipo IL4RA I50V/Q551R. El riesgo de asma estaba notablemente incrementado (P = .007) para la interacción entre IL4RA V50/Q551 (OR, 2.48; 95% CI, 1.25-4.99), mientras por el contrario estaba disminuido (P = .0037) en la interacción entre IL4RA V50/R551 (OR, 0.31; 95% CI, 0.14-0.71). Ambos genotipos parecen ser un factor de riesgo y un factor de protección, respectivamente, en el desarrollo de un fenotipo asmático en nuestra población.
También analizamos el papel de la interacción genética de los polimorfismos localizados sobre cromosomas diferentes. Estas interacciones fueron estudiadas entre los polimorfismos de IL4RA y IL13, y los polimorfismos IL5 y IL13. No se muestran los datos. No encontramos diferencias estadísticamente significativas en la distribución de frecuencias de ambas interacciones genéticas.
Discusión
En los últimos 10 años se han hecho intensos esfuerzos para determinar el contenido molecular del polen de O. europaea (7-20). Conocer de manera precisa los alérgenos que componen el polen del olivo sería un primer paso fundamental en la comprensión de los mecanismos implicados en el desarrollo de esta polinosis.
La disponibilidad de estos alérgenos purificados facilitaría la determinación de los perfiles de reactividad IgE específica de los pacientes alérgicos al olivo. Un amplio estudio con pruebas serológicas y test cutáneos a alérgicos a O. europaea que padecen asma y/ó rinitis pone de manifiesto la existencia de cuatro alérgenos del polen del olivo que resultarían útiles por su fiabilidad, sensibilidad y especificidad en el diagnóstico de la polinosis (21).
El uso de los distintos alérgenos del polen del olivo también facilitaría conocer la implicación de cada una de estas moléculas en la patogénesis de las diferentes enfermedades relacionadas al polen del olivo. En nuestro trabajo analizamos la respuesta de anticuerpos IgE contra dos alérgenos del polen del olivo, Ole e 2 y Ole e10, que son mayoritarios en nuestra población con un porcentaje de reconocimiento superior al 50%. Nuestros resultados también ponen de manifiesto una fuerte asociación con el asma en los pacientes sensibilizados a Ole e 2 y Ole e 10.No obtuvimos resultados similares con ningún otro alérgeno de O. europaea.
Gracias a las evidencias acumuladas, sabemos que el asma es un desorden genético complejo con un fenotipo heterogéneo y que se atribuye a la interacción entre múltiples genes y factores ambientales (22-26). Por este motivo, nosotros estudiamos la influencia de genes localizados en la región MHC del cromosoma 6 (DRB1, DRB3, DRB4, DRB5 y DRQ1) en algunos de los fenotipos asociados al asma como los niveles de IgE totales, específicos frente al extracto completo del polen del olivo, específicos frente a los alérgenos Ole e 2 y Ole e 10.
Ole e 2 (profilina del olivo) es mayoritario en nuestra población. La frecuencia de reconocimiento de Ole e 2 había sido determinada inicialmente por otros grupos en un 24% (18) mediante la técnica de inmunodetección. El grupo de Martínez y colaboradores (29), sin embargo, describen que la profilina del olivo aislada naturalmente y analizada mediante pruebas cutáneas revela una prevalencia de un 66% en pacientes con alergia al polen del olivo polisensibilizados y de un 50% en pacientes monosensibilizados. Nosotros hemos descrito (21) una frecuencia de reconocimiento del 74.6% en una población de Jaén, usando ELISA y test cutáneos. Una explicación de estas discrepancias podría encontrarse en la pérdida de epítopos conformacionales según la metodología empleada.
Sin embargo, aunque las diferentes técnicas diagnósticas podrían influir en la prevalencia de anticuerpos IgE α-Ole e 2, algunas evidencias sugieren que ciertos factores intrínsecos y/ó ambientales podrían influir también. Se ha observado en estudios preliminares que los alergogramas de alérgicos al polen del olivo de áreas con alta exposición, como Jaén, son notablemente diferentes de aquellas áreas con baja exposición. Con relación a esto, nosotros observamos que la caracterización como alérgenos mayoritarios de Ole e 6 (12) y Ole e 2 (18) sólo se produce en poblaciones donde el olivo se cultiva intensamente. El motivo no es claro, pero el hecho de estar sometidos los pacientes a una alta o baja exposición al polen del olivo, sugiere que la exposición es un factor de riesgo en la respuesta de IgE a determinados alérgenos del polen del olivo.
El análisis de la reactividad de IgE frente a Ole e 2 y Ole e 10 revela que ambos alérgenos están relacionados con el fenotipo asmático. Encontramos una correlación estadísticamente significativa entre la presencia de IgE α-Ole e 2 y el asma, así como con los niveles más altos de IgE total, y al extracto completo del olivo. Esta correlación también se observa entre la sensibilización a Ole e 10 y el asma, así como con los niveles de IgE total, IgE al extracto completo y a Ole e 10. Lo más interesante es que cuando analizábamos de forma combinada estos datos encontrábamos que el riesgo de desarrollar asma era mayor en los pacientes que estaban sensibilizados a ambos alérgenos que en aquéllos sensibilizados a uno de ellos únicamente. También comprobamos que los pacientes con anticuerpos IgE α-Ole e 10 presentaban episodios de asma más severos y durante más días que los pacientes no sensibilizados a Ole e 10. Estos datos sugieren que la existencia de IgE α-Ole e 10 en pacientes con asma producido por O. europaea podría servir para determinar el riesgo de desarrollar la enfermedad de forma severa.
El último paso de nuestro estudio fue analizar la relación entre los antígenos HLA de clase II y la respuesta de anticuerpos IgE α-Ole e 2 y Ole e 10, analizando su posible relación con le fenotipo clínico. El análisis global de los resultados nos mostró que había cuatro antígenos relacionados con dicha respuesta: DR7, DR3, DQ2 y DR2 (15). Los resultados de nuestro estudio muestran que los antígenos HLA-DR7 y HLA-DQ2 son un factor de riesgo para la sensibilización a Ole e 2.
En los pacientes sensibilizados a Ole e 10 el antígeno DR2 (15) está relacionado con la respuesta IgE contra este alérgeno y parece ser un factor de riesgo en la sensibilización específica al mismo. Estos datos sugieren que el control genético de la respuesta de IgE alérgeno específica en la alergia al polen del olivo podría ser importante en la clínica de la enfermedad. Estos resultados están de acuerdo con estudios previos que describen la asociación entre HLA-DRB1*02/DRB5 y el asma en pacientes con alergia al polen de gramíneas (30) y la asociación descrita por Moffatt y sus colaboradores (31) de la relación entre el haplotipo extendido de TNF-α (LTα NcoI*/TNF-308*2/HLA-DRB1*2) y el asma.
Respecto al análisis de polimorfismos podemos decir que todas nuestras frecuencias genotípicas fueron similares, para cada polimorfismo estudiado, a las obtenidas por otros grupos en poblaciones europeas.
Nuestro estudio pone de manifiesto que ambos polimorfismos estudiados del gen de IL13 están asociados con la regulación genética de la alergia al polen del olivo. Mientras que el polimorfismo IL13 C-1112T confiere protección frente al desarrollo de alergia a O.europaea, IL13 R130Q, por su parte, es un importante factor de riesgo en el desarrollo de alergia al polen del olivo.
El análisis combinado de SNPs nos muestra una fuerte asociación entre los polimorfismos IL4RA I50V y Q551R y el asma, poniendo de manifiesto la manera en que la acción conjunta de los polimorfismos de varios genes actúa sobre el desarrollo de un fenotipo clínico determinado, mientras que la aparición en solitario de los mismos no tienen ningún efecto.
Aunque IL5 es importante en la inflamación alérgica, ningún estudio ha encontrado asociación entre el polimorfismo de IL5 y el asma per se o parámetros clínicos del asma. Algunos estudios sugieren la posibilidad de que el polimorfismo de IL5 este asociado con la severidad del asma pero no con su desarrollo (36). Nuestros resultados muestran que IL5 no está asociado con el desarrollo de la alergia al polen del olivo, pero sin embargo, el polimorfismo IL5 C-746T parece tener un efecto protector sobre el desarrollo de un fenotipo de asmático en nuestra población. Estos datos son los primeros que asocian un determinado genotipo del polimorfismo IL5 C-746T con el asma.
Varios estudios demuestran que las variaciones genéticas en la estructura de los codones 16 y 27 del receptor b2 adrenérgico altera su función in vitro y está asociado con la severidad del asma y la hiperrespuesta de las vías respiratorias, aunque no con un diagnóstico de asma. En nuestro estudio no hemos encontrado ninguna asociación de estas variantes del receptor b2 adrenérgico y el riesgo de desarrollar alergia al polen del olivo y asma. Encontramos como otros investigadores, que R16 segrega más comúnmente con Q27 y G16 con E27. Estos alelos están en un fuerte desequilibrio de ligamiento y el haplotipos R16E27 es muy poco frecuente (37).
Bibliografía
Bousquet J, Cour P, Guerin B, Michel FB. Allergy in the Mediterranean area I. Pollen counts and pollinosis of Montpellier. Clin Allergy 1985;14:249–258.
Gonzalez EM, Villalba M, Rodrıguez R. Allergenic cross-reactivity of olive pollen. Allergy 2000;55:658–663.
Liccardi G, D’Amato M, D’Amato G. Oleaceae pollinosis: a review. Int Arch Allergy Immunol 1996;111:210–217.
Davies RR, Smith IP. Forecasting the start and severity of the hay fever season. Clin Allergy 1973;3:263–267.
Florido JF, Gonzalez P, Saenz de San Pedro B, Quiralte J, Arias de Saavedra JM, Peralta V et al. High levels of Olea europaea pollen and relation with clinical findings. Int Arch Allergy Immunol 1999;119:133–137.
Rodrıguez R, Villalba M, Monsalve RI, Batanero E. The spectrum of olive pollen allergens. Int Arch Allergy Immunol 2001;125:185–195.
Blanca M, Boulton P, Brostoff J, Gonzalez-Reguera I. Studies of the allergens of Olea europaea pollen. Clin Allergy 1983;13:473–478.
Villalba M, Batanero E, Lopez-Otın C, Sanchez LM, Monsalve RI, Gonzalez de la Peña MA et al. Amino acid sequence of Ole e I, the major allergen from olive tree pollen (Olea europaea). Eur J Biochem 1993;216:863–869.
Lauzurica P, Gurbindo C, Maruri N, Galocha B, Dıaz R, Gonzalez J et al. Olive (Olea europaea) pollen allergens – I. Immunochemical characterization by immunoblotting, CRIE and immunodetection by a monoclonal antibody. Mol Immunol 1988;25:329–335.
Rodrıguez R, Villalba M, Batanero E, Gonzalez EM, Monsalve RI, Huecas S et al. Allergenic diversity of the olive pollen. Allergy 2002;57(Suppl. 71):6–16.
Boluda L, Alonso C, Fernandez-Caldas E. Purification, characterization, and partial sequencing of two new allergens of Olea europaea. J Allergy Clin Immunol 1998;101:210–216.
Batanero E, Ledesma A, Villalba M, Rodrıguez R. Purification, amino acid sequence and immunological characterization of Ole e 6, a cysteine-enriched allergen from olive tree pollen. FEBS Lett 1997;410:293–296.
Batanero E, Villalba M, Ledesma A, Puente XS, Rodrı´ guez R. Ole e 3, an olive-tree allergen, belongs to a widespread family of pollen proteins. Eur J Biochem 1996;241:772–778.
Ledesma A, Villalba M, Rodrıguez R. Cloning, expression and characterization of a novel EF-hand Ca2+-binding protein from olive pollen with allergenic activity. FEBS Lett 2000;466:192–196.
Tejera ML, Batanero E, Villalba M, Rodrıguez R. Identification, isolation and characterization of Ole e 7, a new allergen of olive tree pollen. J Allergy Clin Immunol 1999;104:797–802.
Florido Lopez JF, Quiralte Enriquez J, Arias de Saavedra Alias JM, Saenz de San Pedro B, Martin Casanez E. An allergen from Olea europaea pollen (Ole e 7) is associated with plant-derived food anaphylaxis. Allergy 2002; 57(Suppl. 71):53–59.
Huecas S, Villalba M, Rodrıguez R. Ole e 9, a major olive pollen allergen is a 1,3-b-glucanase. Isolation, characterization, amino acid sequence, and tissue specificity. J Biol Chem 2001;276: 27959–27966.
Ledesma A, Rodrıguez R, Villalba M. Olive pollen profilin. Molecular and immunologic properties. Allergy 1998;53:520–526.
Asturias JA, Arilla MC, Gomez-Bayon N, Martınez J, Martınez A, Palacios R. Cloning and expression of the panallergen profilin and the major allergen (Ole e 1) from olive tree pollen. J Allergy Clin Immunol 1997;100:365–372.
Barral P, Batanero E, Palomares O, Quiralte J, Villalba M, Rodrıguez R. A major allergen from pollen defines a novel family of plant proteins and show intra and inter-species cross-reactivity. J Immunol 2004;172:3644–3651.
Quiralte J, Florido F, Arias de Saavedra JM, Gomez A, Saenz de San Pedro B, Gonzalez E et al. Olive allergen-specific IgE responses in patients with Olea europaea pollinosis. Allergy 2002;57(Suppl. 71):47–52.
Howard TD, Meyers DA, Bleecker ER. Mapping susceptibility genes for allergic diseases. Chest 2003;123(Suppl. 3):363S–368S.
Hakonarson H, Halapi E. Genetic analyses in asthma: current concepts and future directions. Am J Pharmacogenom 2002;2:155–166.
Cardaba B, Vilches C, Martın E, de Andres B, del Pozo V, Hernandez D et al. DR7 and DQ2 are positively associated with immunoglobulin-E response to the main antigen of olive pollen (Ole e I) in allergic patients. Hum Immunol 1993;38:293–299.
Cardaba B, Cortegano I, Florido F, Arrieta I, Aceituno E, de Pozo V et al. Genetic restrictions in olive pollen allergy. J Allergy Clin Immunol 2000;105:292–298.
Cardaba B, Cortegano I, Florido F, Civantos E, del Pozo V, Gallardo S et al. Update in the understanding of genetic predisposition to olive pollen sensitization. Allergy 2002;57(Suppl. 71):41–46.
Dreborg S. Skin tests used in type I allergy testing (position paper). Allergy 1989;44(Suppl. 10):22–31.
Miller SA, Dykes D, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res 1988;16:1215–1218.
Martınez A, Asturias JA, Monteseirin J, Moreno V, García-Cubillana A, Herna ndez M et al. The allergenic relevance of profilin (Ole e 2) from Olea europaea pollen. Allergy 2002;57(Suppl. 71):17– 23.
Woszczek G, Kowalski ML, Borowiec M. Association of asthma and total IgE levels with human leucocyte antigen-DR in patients with grass allergy. Eur Respir J 2002;20:79–85.
Moffatt MF, James A, Ryan G, Musk AW, Cookson WO. Extended tumour necrosis factor/HLA-DR haplotypes and asthma in an Australian population sample. Thorax 1999;54:757–761.
Graves PE, Kabesch M, Halonen M, Holberg CJ, Baldini M, Fritzsch C, Weiland SK, Erickson RP, von Mutius E, Martinez FD. A cluster of seven tightly linked polymorphisms in the IL-13 gene is associated with total serum IgE levels in three populations of white children. J Allergy Clin Immunol. 2000 Mar;105(3):506-13.
Noguchi E, Shibasaki M, Arinami T, Takeda K, Yokouchi Y, Kobayashi K, Imoto N, Nakahara S, Matsui A, Hamaguchi H. No association between atopy/asthma and the ILe50Val polymorphism of IL-4 receptor. Am J Respir Crit Care Med. 1999 Jul;160(1):342-5.
Large V, Hellstrom L, Reynisdottir S, Lonnqvist F, Eriksson P, Lannfelt L, Arner P. Human beta-2 adrenoceptor gene polymorphisms are highly frequent in obesity and associate with altered adipocyte beta-2 adrenoceptor function. J Clin Invest. 1997 Dec 15;100(12):3005-13.
Deichmann KA, Schmidt A, Heinzmann A, Kruse S, Forster J, Kuehr J. Association studies on beta2-adrenoceptor polymorphisms and enhanced IgE responsiveness in an atopic population. Clin Exp Allergy. 1999 Jun;29(6):794-9.
Hong SJ, Lee SY, Kim HB, Kim JH, Kim BS, Choi SO, Lee SG, Shin ES, Hong TJ. IL-5 and thromboxane A2 receptor gene polymorphisms are associated with decreased pulmonary function in Korean children with atopic asthma. J Allergy Clin Immunol. 2005 Apr;115(4):758-63.
Turner SW, Khoo SK, Laing IA, Palmer LJ, Gibson NA, Rye P, Landau LI, Goldblatt J, Le Souef PN. beta2 adrenoceptor Arg16Gly polymorphism, airway responsiveness, lung function and asthma in infants and children. Clin Exp Allergy. 2004 Jul;34(7):1043-8.
{kind=link}
{kind=link}